
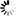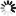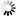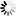 loading subjects...
loading subjects...

To elucidate the basis for the drop in pressure from large artery to vein, Jean Léonard Marie Poiseuille (1797-1869) undertook a series of seminal studies that defined the laws of fluid flow in tubes of uniform cross-section and that identified the microcirculation as the major site of the resistance to flow [1, 2]. While best known for his meticulous experimental studies of viscous flow in glass tubes [2], he also made numerous observations in the mesentery of the frog and other microvascular preparations; these observations served to highlight the dynamics of red cell distribution, the presence of the annular plasma layer and the “skimming” of plasma by capillaries, and the adhesion of white cells to the endothelium of post-capillary venules [3]. Yet it wasn't until almost a century later that physiologists began to methodically explore the resistance to flow within the microcirculation proper using techniques of intravital microscopy. Landis [4] pioneered many quantitative methods for describing microvascular structure and function. Using finely drawn pipettes inserted into microvessels of frog mesentery, and calculating the velocity of bolus infusions of dyes through successive microvascular divisions, Landis attempted to calculate the resistance to flow within the capillary network and concluded that Poiseulle's law cannot be applied to the flow of blood through the capillary network except in a very limited sense. However, with the advent of sophisticated instruments for measurement of capillary pressure [5, 6] and red blood cell velocity [7], relatively precise quantitative measurements of pressure drops [8] and flow rates [9, 10] could be obtained; such measurements thus provided direct in situ flow resistance data for the hierarchy of microvessels from arteriole to venule [11].
As shown in Figure 1, measurements of pressure drops and flows in single unbranched microvessels [11] reveal that over the broad range of diameters (D) within the microcirculation proper, Poiseuille's fourth power relationship (i.e., resistance varies inversely with D4) far overshadows other determinants of resistance in the normal flow state. The significance of this relationship cannot be understated in light of the fact that control and regulation of microvascular blood flow is manifest by the ability of the vascular system to alter resistance in response to vasomotor adjustments. However, striking departures from Poiseuille's relationship may dictate the outcome of pathological flow states, with some examples including the low flow state, inflammation, and blood cell disorders. While the resistance to flow spans nearly five decades as blood courses its way from arteriole to venule, the large scatter in the experimental data may reflect significant departures from the flow of a Newtonian fluid through a smooth walled tube of circular cross-section. The effects of irregularities in geometry, broad variations in microvessel hematocrit and shear rates, blood cell deformability, red cell aggregation and blood cell adhesion to the endothelium are reviewed in the following.